Durante mucho tiempo, cuando alguien mencionaba el lactato, automáticamente pensábamos en dolor muscular, fatiga o en ese “ardor” que sentimos al final de una serie intensa. Pero ¿y si te dijera que esa percepción está completamente desactualizada? Hoy, gracias a décadas de investigación, sabemos que el lactato es mucho más que un mal llamado residuo del metabolismo: es una molécula fundamental para nuestra fisiología, una señal que conecta tejidos, regula genes y mejora el rendimiento.
Durante más de un siglo, el lactato fue considerado un residuo metabólico generado por la falta de oxígeno en el ejercicio intenso. Esta visión, nacida en los experimentos de Otto Meyerhof y A.V. Hill a principios del siglo XX, sostenía que el lactato causaba fatiga, acidosis y bajo rendimiento.
Sin embargo, la revolución conceptual que ha tenido lugar en los últimos 30 años, liderada por autores como George A. Brooks, ha cambiado radicalmente esta perspectiva. Hoy sabemos que el lactato no solo no es un producto de desecho, sino que es una molécula clave en la regulación energética, en la señalización metabólica y en procesos adaptativos sistémicos inducidos por el ejercicio físico, que redefine al lactato como una mióquina y exerkina: es decir, una molécula producida por el músculo y liberada durante el ejercicio con efectos locales y sistémicos sobre el metabolismo, la inflamación, la neuroplasticidad y la señalización génica.
El lactato, entonces, no solo transporta energía, sino que también transmite información entre tejidos y órganos durante y después del ejercicio.
Este artículo de blog sintetiza los aspectos fundamentales sobre esta revisión realizada por Brooks et al 2023 que sintetiza esta visión contemporánea del lactato como una molécula señalizadora multifuncional, en donde se trata de indagar sobre ¿qué implica realmente que el lactato sea una señal fisiológica? ¿Qué rutas modula? ¿Y por qué su redescubrimiento puede cambiar la forma en que entendemos el metabolismo, la salud y la enfermedad?
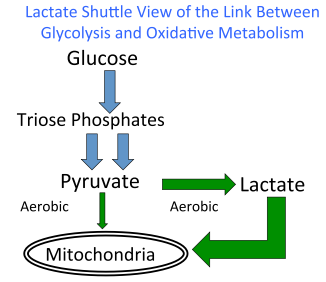
Durante mucho tiempo, la aparición de lactato fue asociada casi exclusivamente a situaciones de deficiencia de oxígeno, como si su presencia en el organismo fuese prueba irrefutable de un metabolismo anaeróbico en acción. Sin embargo, múltiples líneas de evidencia provenientes de estudios en animales intactos, preparaciones de tejido y ensayos en humanos han desmontado esta idea, demostrando con solidez que la glucólisis —el proceso que transforma glucosa y glucógeno en energía— conduce a la producción de lactato incluso bajo condiciones plenamente aeróbicas.
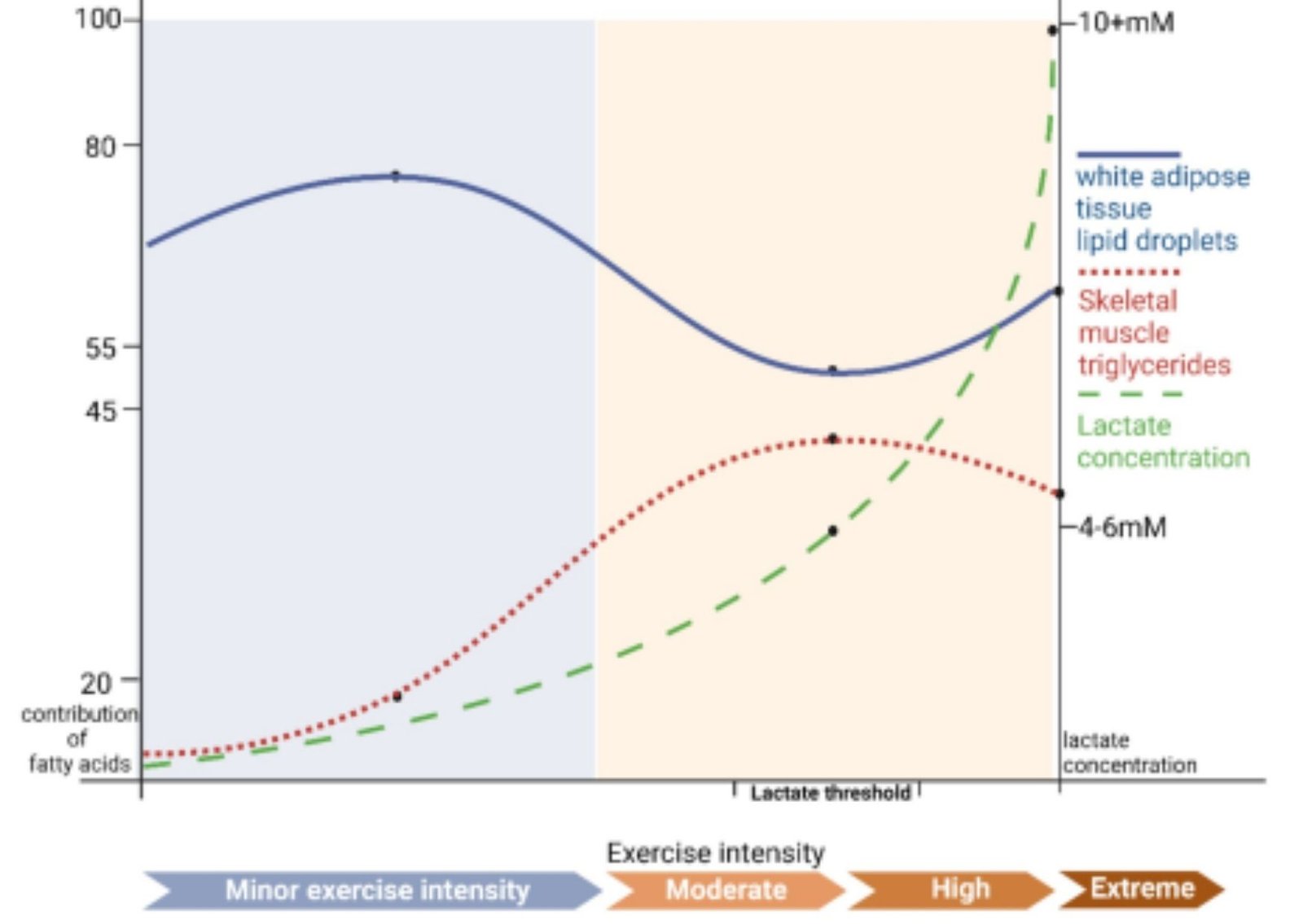
En humanos sanos en reposo, tanto el músculo esquelético como la sangre arterial presentan concentraciones estables de lactato que rondan 1,0 mM, mientras que el piruvato circulante suele encontrarse en torno a los 0,1 mM. Esta proporción da lugar a una relación lactato/piruvato (L/P) de aproximadamente 10, un valor indicativo de que la conversión de piruvato a lactato ocurre activamente aun cuando el oxígeno está abundantemente disponible. Esta transformación no se debe a una falta de oxígeno, sino a la dinámica del metabolismo celular y al mantenimiento del equilibrio redox citosólico.
De hecho, la presión parcial de oxígeno (PO₂) intramuscular en reposo alcanza valores cercanos a los 40 Torr, considerablemente por encima del umbral crítico necesario para la respiración mitocondrial eficiente, estimado entre 1 y 2 Torr. Por lo tanto, la producción y liberación de lactato en estas condiciones no obedece a un déficit de oxígeno, sino a la propia organización funcional del metabolismo energético en tejidos bien oxigenados .
Durante el ejercicio, este fenómeno se acentúa. A intensidades que alcanzan aproximadamente el 65 % del consumo máximo de oxígeno (VO₂máx.), los músculos activos incrementan significativamente tanto la producción como la liberación de lactato. A pesar de este aumento, la PO₂ intramuscular se mantiene en torno a los 3–4 Torr, aún por encima del umbral mitocondrial crítico. Paralelamente, la relación L/P en sangre puede elevarse más de un orden de magnitud, alcanzando valores de hasta 500, lo que refleja un cambio profundo en el estado redox celular, sin que esto implique hipoxia en el tejido.
En este contexto, resulta crucial destacar que la mayor parte del lactato generado —entre un 75 % y un 80 %— es rápidamente reutilizado, ya sea dentro del mismo músculo que lo produce o tras ser captado por otros tejidos. El corazón, por ejemplo, lo oxida de forma eficiente como fuente energética, mientras que el hígado lo emplea como sustrato para la gluconeogénesis, cerrando así el ciclo de reciclaje del carbono energético.
En conjunto, estos hallazgos confirman que la glucólisis hasta lactato no es sinónimo de metabolismo anaeróbico, y que la presencia de lactato debe entenderse como parte integral del metabolismo aeróbico normal. Esta reinterpretación no solo desmonta viejas ideas en fisiología, sino que abre nuevas perspectivas para comprender el papel del lactato como molécula señalizadora y reguladora del metabolismo sistémico
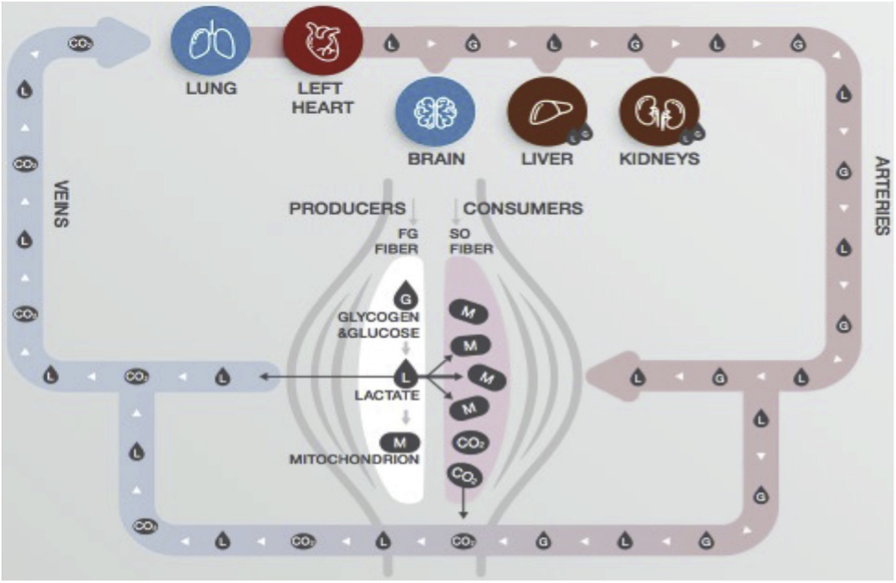
Una de las contribuciones clave del trabajo de Brooks es el concepto de lactate shuttle, o transporte de lactato, una red de comunicación metabólica entre diferentes órganos y tejidos. En este modelo, el lactato no es un residuo sino un vector de información y energía que conecta al músculo con el cerebro, el corazón, el hígado, el tejido adiposo e incluso el intestino. Este transporte ocurre tanto en condiciones de reposo —por ejemplo, tras una comida rica en carbohidratos— como durante el ejercicio intenso.
Así, durante el esfuerzo físico, las fibras musculares tipo II, de metabolismo glucolítico, generan grandes cantidades de lactato. Este lactato es rápidamente exportado al torrente sanguíneo y captado por tejidos oxidativos como el corazón y las fibras tipo I, donde es convertido nuevamente en piruvato y oxidable en la mitocondria. Esta circulación interna de carbono no solo permite el reciclaje energético, sino que lleva consigo un conjunto de señales bioquímicas que modifican la expresión génica, el estado redox celular, y las decisiones metabólicas de las células receptoras.
Pero lo más notable es que estas funciones señalizadoras del lactato no se restringen al ejercicio. También están presentes en condiciones fisiológicas tan diversas como la digestión, la neurogénesis, el desarrollo embrionario, el sueño, y la reparación tisular. Este hecho refuerza la idea de que el lactato no es un simple “intermediario metabólico”, sino una verdadera “LACTOHORMONA”, como lo denominó el propio Brooks en publicaciones anteriores.
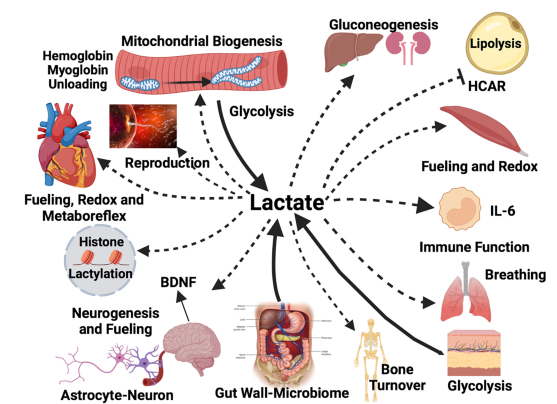
En la imagen se observan diversas formas de transporte intracelular de lactato. Las células y tejidos productores de lactato (impulsores) (líneas continuas y puntas de flecha) que contribuyen al lactato circulante incluyen contribuciones del tegumento, el intestino, el fibras musculares de contracción rápida, fibras musculares lentas posprandial y fibras musculares mixtas al inicio del ejercicio. Los receptores de lactato que lo eliminan (líneas discontinuas y puntas de flecha menores) incluyen la oxidación mitocondrial del lactato en el músculo esquelético rojo y mixto, el corazón y el cerebro durante el ejercicio a ritmo constante. También se incluyen (líneas discontinuas y puntas de flecha menores) la eliminación de lactato mediante gluconeogénesis en el hígado y los riñones, y en las neuronas cerebrales (como parte del ANLS). La liberación de IL-6 estimulada por lactato desde los monocitos y el músculo activo es un ejemplo de liberación de citocinas estimulada por lactato. Ya sean impulsoras o receptoras, todas las células experimentan efectos de señalización redox. Los sitios de señalización que no implican intercambio ni transformación de carbono incluyen el tejido adiposo blanco, donde el lactato inhibe la lipólisis mediante la señalización de HCAR y CREB; el corazón, donde la acumulación de lactato en el músculo periférico estimula el metaborreflejo con señalización aferente al centro cardiovascular medular a través de fibras sensoriales de tipo III y IV, lo que aumenta el gasto cardíaco; la ventilación pulmonar a través del receptor olfativo del cuerpo carotídeo (Olfr78); el músculo esquelético, donde estimula la biogénesis mitocondrial a través del coactivador-1 alfa del receptor activado por el proliferador de peroxisomas gamma (PGC-1a); las especies reactivas de oxígeno (ROS) y la activación de las sirtuinas. Además, el lactato tiene las siguientes acciones: en el músculo en funcionamiento, disocia la oximioglobina y la oxihemoglobina sanguínea; En el cerebro, el lactato procedente de la circulación arterial de la glucólisis en los astrocitos alimenta las neuronas y participa en la señalización glutamatérgica, además de estimular la neurogénesis en el hipocampo y la secreción del factor neurotrópico derivado del cerebro (BDNF). Además, la lactatemia y la acumulación tisular de lactato tienen un efecto epigenético a través de la lactilación de histonas, y el lactato tiene efectos antiinflamatorios. Tejidos implicados, comenzando arriba a la izquierda y en sentido horario: fibras musculares esqueléticas, órganos gluconeogénicos (hígado y riñones), tejido adiposo blanco, músculo esquelético rojo activo, monocitos, pulmones, tegumento, esqueleto, pared intestinal y microbioma, cerebro, todas las células nucleadas que contienen ADN, corazón, óvulos y espermatozoides.
Una de las grandes revelaciones del artículo es el reconocimiento del lactato como agente epigenético. Gracias a investigaciones recientes, se ha descubierto que esta molécula puede inducir la lactilación de histonas, un proceso por el cual se añaden grupos lactilo a proteínas nucleares que regulan la expresión génica. Este hallazgo sitúa al lactato en el centro del diálogo entre metabolismo y genética, mostrando que no solo produce energía, sino que instruye a la célula sobre qué genes activar en respuesta a un entorno cambiante.
La señalización mediada por lactato también incluye la activación de vías clásicas como PGC-1α (coactivador del receptor gamma activado por proliferadores de peroxisomas), SIRT1/3, AMPK y ROS, todas ellas conocidas por regular la biogénesis mitocondrial, la adaptación al entrenamiento y el envejecimiento celular. Así, la exposición repetida al lactato —como ocurre en el ejercicio físico regular— puede ser vista como una forma de estimulación hormética que fortalece la resiliencia metabólica de la célula.
Incluso más allá del núcleo, el lactato también modula la función endocrina. Por ejemplo, inhibe la lipólisis en tejido adiposo mediante la activación del receptor HCAR-1, regula la liberación de testosterona en células de Leydig, y participa en el control del apetito post-ejercicio a través de su efecto supresor sobre la hormona ghrelina. Estas acciones endócrinas subrayan el papel del lactato como molécula organizadora de la respuesta sistémica al esfuerzo, adaptando múltiples sistemas al estado energético actual.
Uno de los ejemplos más fascinantes de la versatilidad del lactato se encuentra en su relación con el sistema nervioso. En el cerebro, el lactato no solo actúa como fuente energética —lo que ya es relevante, dado el elevado costo metabólico del tejido nervioso—, sino que modula procesos de plasticidad sináptica y formación de memoria. Durante el ejercicio, el lactato generado por el músculo cruza la barrera hematoencefálica y estimula la liberación de BDNF (factor neurotrófico derivado del cerebro), un componente esencial para la neurogénesis y el aprendizaje.
Además, en el contexto del “astrocyte-neuron lactate shuttle” (ANLS), los astrocitos generan lactato a partir de glucosa y lo transfieren a las neuronas, donde es oxidado para sostener la actividad sináptica. Este sistema no solo asegura la homeostasis energética cerebral, sino que convierte al lactato en una molécula integradora entre metabolismo, neurotransmisión y plasticidad neuronal. De allí que la suplementación con lactato o su infusión intravenosa se haya propuesto como terapia en pacientes con traumatismo craneoencefálico o deterioro cognitivo.
El artículo también reconoce que el papel del lactato no siempre es beneficioso. En ciertas condiciones, como la inflamación crónica o el cáncer, el lactato puede participar en procesos patológicos. En el contexto oncológico, por ejemplo, el lactato producido por las células tumorales favorece la angiogénesis, suprime respuestas inmunes antitumorales y promueve la acidificación del microambiente, facilitando la invasión celular.
Sin embargo, esta misma molécula, en contextos controlados, puede estimular la actividad de linfocitos CD8⁺ y promover respuestas antitumorales, como se ha visto con la administración exógena de lactato en modelos murinos. Esta dualidad sugiere que el lactato no es inherentemente “bueno” o “malo”, sino que su impacto depende del contexto, del tipo celular y del balance entre producción y eliminación. Así como una hormona puede ser beneficiosa o nociva según su concentración y su entorno, el lactato actúa como un regulador contextual.
El redescubrimiento del lactato como señal metabólica nos obliga a repensar categorías fundamentales de la biología. No estamos ante una simple molécula energética, sino ante un verdadero integrador fisiológico: un mensajero bioquímico que informa al cuerpo sobre su estado energético, regula respuestas genéticas, modula la función de órganos distantes y prepara al organismo para adaptarse al entorno.
Desde el entrenamiento deportivo hasta la nutrición, pasando por la inmunología, la neurociencia y la oncología, el lactato aparece como un nodo central en la red de señales que mantienen la salud y la funcionalidad del cuerpo humano. Integrarlo a nuestra comprensión moderna de la fisiología no solo permite optimizar el rendimiento físico y cognitivo, sino también desarrollar nuevas estrategias terapéuticas para enfermedades complejas.
A fin de cuentas, el lactato dejó de ser un subproducto para convertirse en protagonista. Y como toda gran transformación científica, esta revalorización nos recuerda que incluso los conceptos más establecidos pueden cambiar radicalmente cuando se observa con nuevos ojos.
Para leer el articulo original sigue el enlace